
-
Product Name
Anti-PLK1 Rabbit antibody
- Documents
-
Description
PLK1 Rabbit monoclonal antibody
-
Tested applications
WB, ICC/IF, FC, IP
-
Species reactivity
Human
-
Alternative names
PLK; STPK13 antibody
-
Isotype
Rabbit IgG
-
Preparation
Antigen: A synthetic peptide of human PLK1
-
Clonality
Monoclonal
-
Formulation
50nM Tris-Glycine(pH 7.4), 0.15M Nacl, 40%Glycerol, 0.01% sodium azide and 0.05% BSA.
-
Storage instructions
Store at 4°C short term. Store at -20°C long term. Avoid freeze / thaw cycle.
-
Applications
WB: 1/1000
ICC/IF: 1/50
FC: 1/20
IP: 1/20
-
Validations
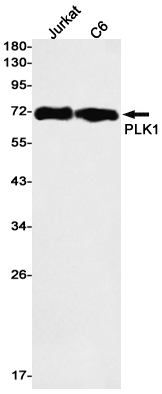
Western blot detection of PLK1 in Jurkat,C6 cell lysates using PLK1 Rabbit mAb(1:1000 diluted).Predicted band size:68kDa.Observed band size:68kDa.
-
Background
Swiss-Prot Acc.P53350.Serine/threonine-protein kinase that performs several important functions throughout M phase of the cell cycle, including the regulation of centrosome maturation and spindle assembly, the removal of cohesins from chromosome arms, the inactivation of anaphase-promoting complex/cyclosome (APC/C) inhibitors, and the regulation of mitotic exit and cytokinesis. Polo-like kinase proteins acts by binding and phosphorylating proteins are that already phosphorylated on a specific motif recognized by the POLO box domains. Phosphorylates BORA, BUB1B/BUBR1, CCNB1, CDC25C, CEP55, ECT2, ERCC6L, FBXO5/EMI1, FOXM1, KIF20A/MKLP2, CENPU, NEDD1, NINL, NPM1, NUDC, PKMYT1/MYT1, KIZ, PPP1R12A/MYPT1, PRC1, RACGAP1/CYK4, SGO1, STAG2/SA2, TEX14, TOPORS, p73/TP73, TPT1, WEE1 and HNRNPU. Plays a key role in centrosome functions and the assembly of bipolar spindles by phosphorylating KIZ, NEDD1 and NINL. NEDD1 phosphorylation promotes subsequent targeting of the gamma-tubulin ring complex (gTuRC) to the centrosome, an important step for spindle formation. Phosphorylation of NINL component of the centrosome leads to NINL dissociation from other centrosomal proteins. Involved in mitosis exit and cytokinesis by phosphorylating CEP55, ECT2, KIF20A/MKLP2, CENPU, PRC1 and RACGAP1. Recruited at the central spindle by phosphorylating and docking PRC1 and KIF20A/MKLP2; creates its own docking sites on PRC1 and KIF20A/MKLP2 by mediating phosphorylation of sites subsequently recognized by the POLO box domains. Phosphorylates RACGAP1, thereby creating a docking site for the Rho GTP exchange factor ECT2 that is essential for the cleavage furrow formation. Promotes the central spindle recruitment of ECT2. Plays a central role in G2/M transition of mitotic cell cycle by phosphorylating CCNB1, CDC25C, FOXM1, CENPU, PKMYT1/MYT1, PPP1R12A/MYPT1 and WEE1. Part of a regulatory circuit that promotes the activation of CDK1 by phosphorylating the positive regulator CDC25C and inhibiting the negative regulators WEE1 and PKMYT1/MYT1. Also acts by mediating phosphorylation of cyclin-B1 (CCNB1) on centrosomes in prophase. Phosphorylates FOXM1, a key mitotic transcription regulator, leading to enhance FOXM1 transcriptional activity. Involved in kinetochore functions and sister chromatid cohesion by phosphorylating BUB1B/BUBR1, FBXO5/EMI1 and STAG2/SA2. PLK1 is high on non-attached kinetochores suggesting a role of PLK1 in kinetochore attachment or in spindle assembly checkpoint (SAC) regulation. Required for kinetochore localization of BUB1B. Regulates the dissociation of cohesin from chromosomes by phosphorylating cohesin subunits such as STAG2/SA2. Phosphorylates SGO1: required for spindle pole localization of isoform 3 of SGO1 and plays a role in regulating its centriole cohesion function. Mediates phosphorylation of FBXO5/EMI1, a negative regulator of the APC/C complex during prophase, leading to FBXO5/EMI1 ubiquitination and degradation by the proteasome. Acts as a negative regulator of p53 family members: phosphorylates TOPORS, leading to inhibit the sumoylation of p53/TP53 and simultaneously enhance the ubiquitination and subsequent degradation of p53/TP53. Phosphorylates the transactivation domain of the transcription factor p73/TP73, leading to inhibit p73/TP73-mediated transcriptional activation and pro-apoptotic functions. Phosphorylates BORA, and thereby promotes the degradation of BORA. Contributes to the regulation of AURKA function. Also required for recovery after DNA damage checkpoint and entry into mitosis. Phosphorylates MISP, leading to stabilization of cortical and astral microtubule attachments required for proper spindle positioning (PubMed:8991084, PubMed:11202906, PubMed:12207013, PubMed:12447691, PubMed:12524548, PubMed:12738781, PubMed:12852856, PubMed:12939256, PubMed:14532005, PubMed:14734534, PubMed:15070733, PubMed:15148369, PubMed:15469984, PubMed:16198290, PubMed:16247472, PubMed:16980960, PubMed:17081991, PubMed:17351640, PubMed:17376779, PubMed:17617734, PubMed:18174154, PubMed:18331714, PubMed:18418051, PubMed:18477460, PubMed:18521620, PubMed:18615013, PubMed:19160488, PubMed:19351716, PubMed:19468300, PubMed:19468302, PubMed:19473992, PubMed:19509060, PubMed:19597481, PubMed:23455478, PubMed:23509069). Together with MEIKIN, acts as a regulator of kinetochore function during meiosis I: required both for mono-orientation of kinetochores on sister chromosomes and protection of centromeric cohesin from separase-mediated cleavage . Phosphorylates CEP68 and is required for its degradation (PubMed:25503564). Regulates nuclear envelope breakdown during prophase by phosphorylating DCTN1 resulting in its localization in the nuclear envelope (PubMed:20679239). Phosphorylates the heat shock transcription factor HSF1, promoting HSF1 nuclear translocation upon heat shock (PubMed:15661742). Phosphorylates HSF1 also in the early mitotic period; this phosphorylation regulates HSF1 localization to the spindle pole, the recruitment of the SCF(BTRC) ubiquitin ligase complex induicing HSF1 degradation, and hence mitotic progression (PubMed:18794143). Regulates mitotic progression by phosphorylating RIOK2 (PubMed:21880710).
Related Products / Services
Please note: All products are "FOR RESEARCH USE ONLY AND ARE NOT INTENDED FOR DIAGNOSTIC OR THERAPEUTIC USE"